Wspomnieliśmy, że Phiomia i Palaeomastodon nie są zaliczane do właściwych słoniokształtnych min. z tego powodu, że w uzębieniu dorosłych osobników współwystępują zęby trzonowe i przedtrzonowe. O co tu chodzi? Dlaczego ma to znaczenie? Otóż za ważną cechę, charakteryzującą i łączącą ze sobą prawdziwe słoniokształtne, w tym współczesne słonie, jest tzw. horyzontalna wymiana uzębienia.
U większości ssaków, w tym u nas – ludzi, najpierw wyżyna się komplet zębów mlecznych, który zwykle obejmuje siekacze, kły i zęby przedtrzonowe. Ten pierwszy zestaw z czasem wypada i jest zastępowany przez garnitur zębów stałych, poszerzony o wyżynające się dopiero teraz zęby trzonowe. Zęby trzonowe nie mają swoich mlecznych prekursorów!
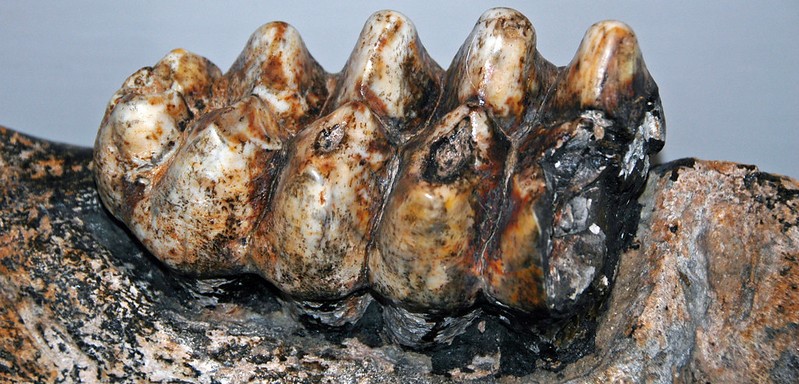
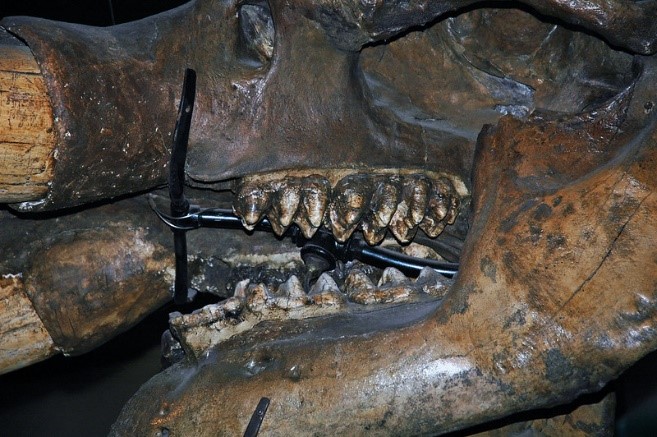
Otóż w szczękach współczesnych słoni, jak i kopalnych mamutów, zazwyczaj występuje maksymalnie do 2-3 zębów policzkowych jednocześnie w każdej gałęzi żuchwy, czy odpowiednio górnej szczęki. Taka sytuacja może mieć miejsce u młodocianych osobników. Dorosłe osobniki mają zazwyczaj tylko jeden, ewentualnie dwa zęby w każdej gałęzi.
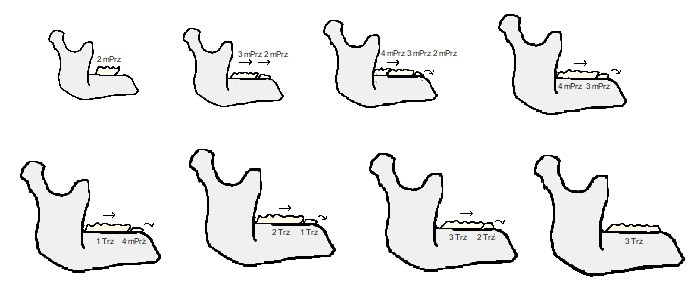
Polega to na tym (ryc. 13), że zęby słoni nie wyżynają się wszystkie naraz, ale po kolei. Uważa się, że najpierw pojawia się jeden, mleczny ząb przedtrzonowy. W miarę jak zwierzę rośnie, za tym zębem, z tyłu, zaczyna wyżynać się kolejny, ale jednocześnie ten pierwszy ściera się w wyniku spożywania przez zwierzę twardej roślinności. Tutaj dzieje się coś niezwykłego.
W wyniku zmian zachodzących w żuchwie (i tak samo w szczęce górnej), tylny, nowy ząb jest stopniowo przesuwany ku przodowi, tak, że w końcu zastępuje starty, pierwszy ząb, którego „ogryzek” w końcu wypada. Za drugim z zębów pojawia się z kolei trzeci mleczny ząb przedtrzonowy i proces się powtarza. A potem znowu, z tym tylko, że kolejny pojawiający się ząb, zdaniem badaczy, odpowiada już pierwszemu zębowi trzonowemu, a nie stałemu przedtrzonowcowi.
Za tym zębem potem pojawiają się jeszcze dwa kolejne, niemniej w jednym czasie funkcjonalny jest zwykle tylko jeden trzonowiec, któremu ewentualnie przez pewien czas może towarzyszyć „ogryzek” poprzednika. W ten sposób słonie, w ciągu swojego życia, mają w sumie 6 par zębów ciernych, odpowiednio w górnej i dolnej szczęce – drugi mleczny przedtrzonowiec, trzeci mleczny przedtrzonowiec, czwarty mleczny przedtrzonowiec, pierwszy trzonowiec, drugi trzonowiec i trzeci trzonowiec. Nigdy jednak nie występują one wszystkie razem.
Pierwsze słoniokształtne miały nieco bogatsze uzębienie. Wczesne formy często wciąż miały stałe przedtrzonowce, zastępujące zęby mleczne. Zęby te były jednak wkomponowane w nowy system horyzontalnego przesuwania się i wymiany zębów trzonowych, które wyżynały się za nimi. Ten mechanizm nie był jeszcze wykształcony u Phiomia i Palaeomastodon. Wszystkie prawdziwe słoniokształtne jednak ten mechanizm już posiadały. Nie jest jednak jasne, czy powstał on tylko raz w ewolucji tej grupy, czy być może do jego wykształcenia się doszło niezależnie w przynajmniej dwóch liniach ewolucyjnych.
Podobnie, jak to miało miejsce z dinoteridami, słoniokształtne przeszły okres ogromnego rozkwitu w neogenie, tj. począwszy mniej więcej od 23 mln. lat temu. Szybko podzieliły się one na dwie duże grupy – „mastodonty” (rozumiane tu jako rodzina Mammutidae i jej najbliżsi krewni) i Elephantida.
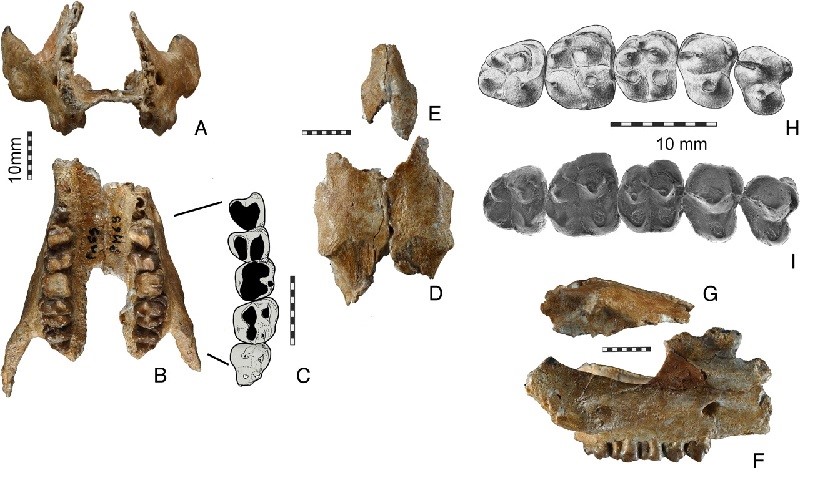
Najstarszym znanym nam mastodontem był Eozygodon morotoensis, pochodzący z wczesno-mioceńskich utworów Kenii, Ugandy i Namibii. Mastodonty szybko jednak rozprzestrzeniły się na całą Eurazję, by następnie przedostać się do Ameryki Północnej. Z Ameryki północnej pochodzi najlepiej znany mastodont – mastodont amerykański (Mammut americanum). Zamieszkiwał on cieplejsze, wciąż zalesione rejony kontynentu w czasie plejstocenu, tj. epoki lodowcowej i wyginął wraz z jej końcem, ok. 10 000 lat temu.









{kind=link}