od lasu po stepo-tundrę
Od lasu po stepo-tundrę ewolucja mamutów
Większości z nas nazwa „mamut” kojarzy się dość jednoznacznie. Przed oczami staje nam obraz włochatego słonia o brunatnym futrze i małych uszach, doskonale przystosowanego do życia w mroźnym klimacie ostatniego zlodowacenia. Gatunek, który sobie wyobrażamy, to mamut włochaty (Mammuthus primigenius). Zwierzę to nie wzięło się jednak znikąd – tak, jak wszystkie inne organizmy żywe na naszej planecie, było ono wynikiem ewolucji. W przeszłości geologicznej żyły też inne mamuty, tj. słoniowate, które zalicza się do rodzaju Mammuthus. O zwierzętach tych wspomnieliśmy w naszym artykule poświęconym ewolucji trąbowców – szerszej grupy, w skład której wchodzą również słoniowate. W niniejszym opracowaniu chcemy czytelnikowi przybliżyć dzieje samej tylko mamuciej linii ewolucyjnej.
Z punktu widzenia nowoczesnej systematyki organizmów, opartej o pokrewieństwa ewolucyjne, rodzaj Mammuthus łączy w sobie wszystkie te gatunki słoniowatych, które były spokrewnione bliżej z mamutem włochatym i ze sobą nawzajem, niż z jakimkolwiek innym słoniem (kopalnym lub współczesnym). W paleontologii rzadko jednak możemy określać stopień pokrewieństwa między zwierzętami na podstawie badań genetycznych, tak jak coraz częściej robi się to z organizmami współczesnymi. Zabieg taki jest możliwy głównie w stosunku do najpóźniejszych mamutów, których zakonserwowane w wieloletniej zmarzlinie szczątki zachowały się na Syberii i Alasce. Dzięki takim badaniom wiemy na przykład (ryc. 1), że najbliższym współczesnym krewniakiem mamutów jest słoń azjatycki (który jest jednak zaliczany do odrębnego rodzaju: Elephas).
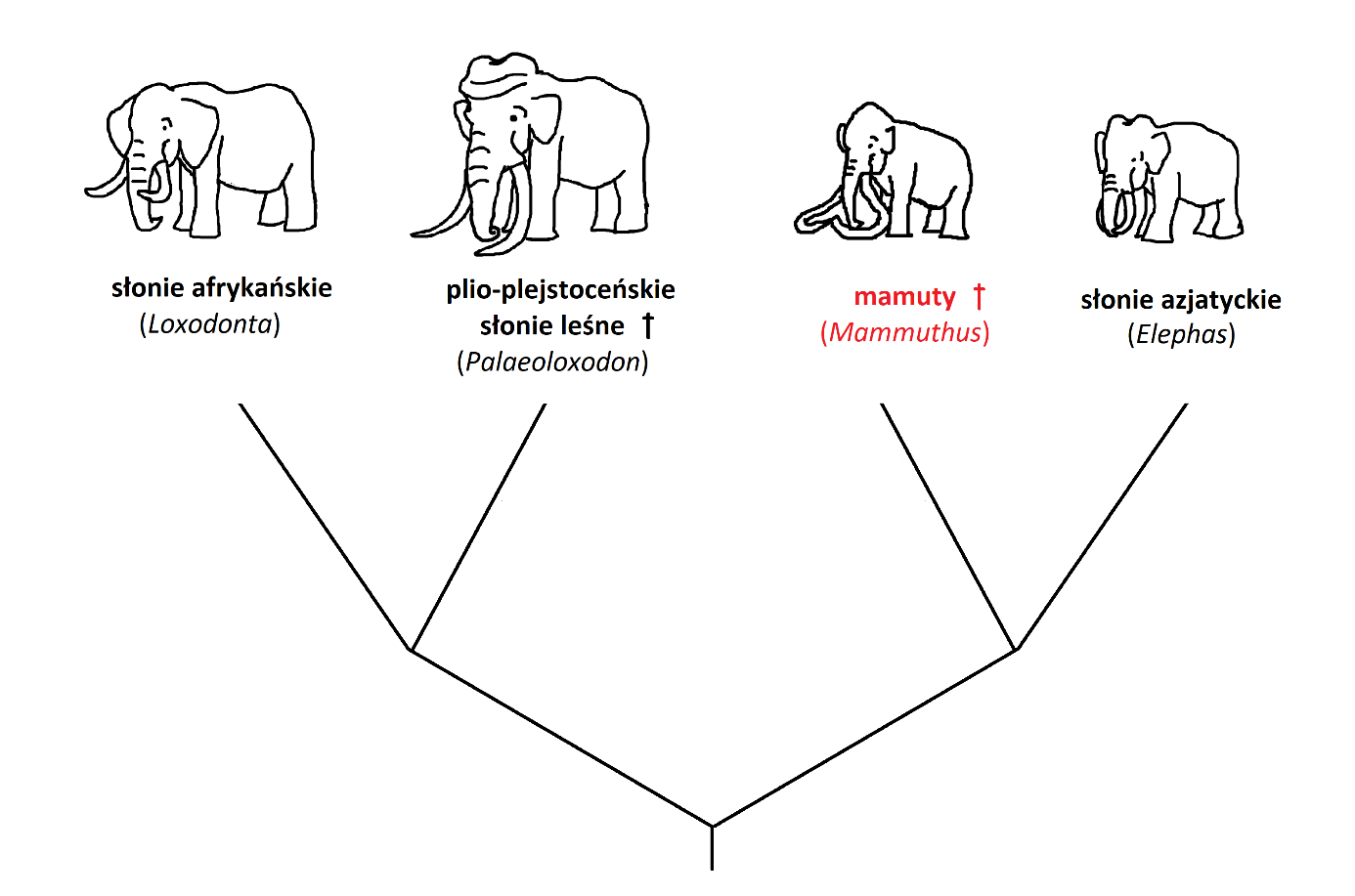
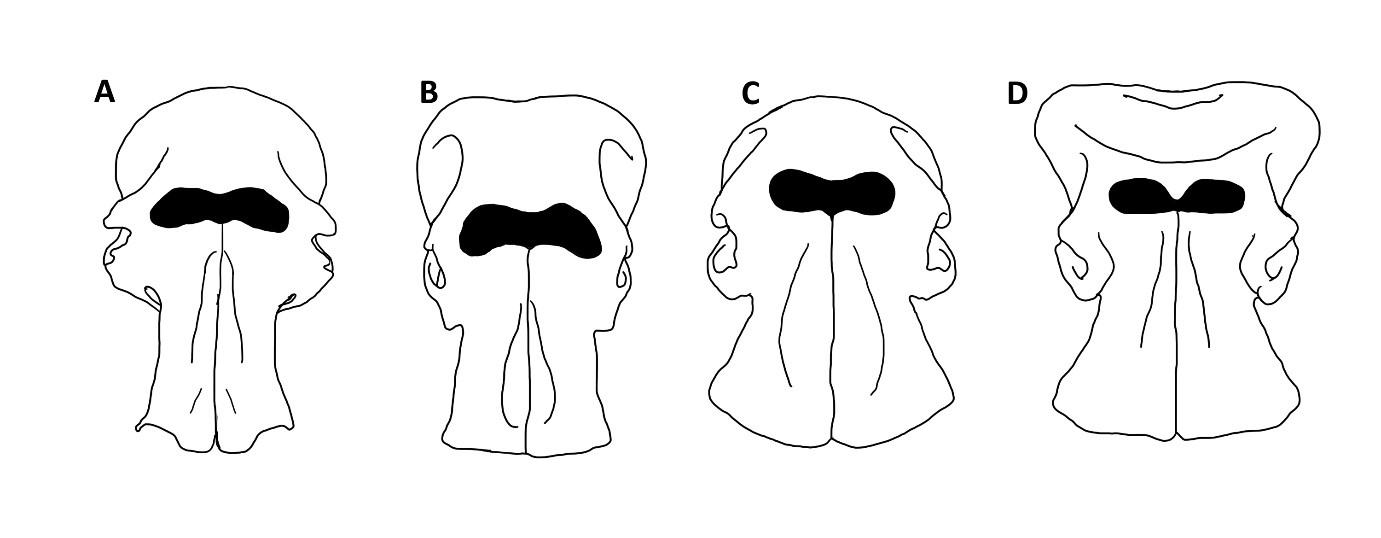
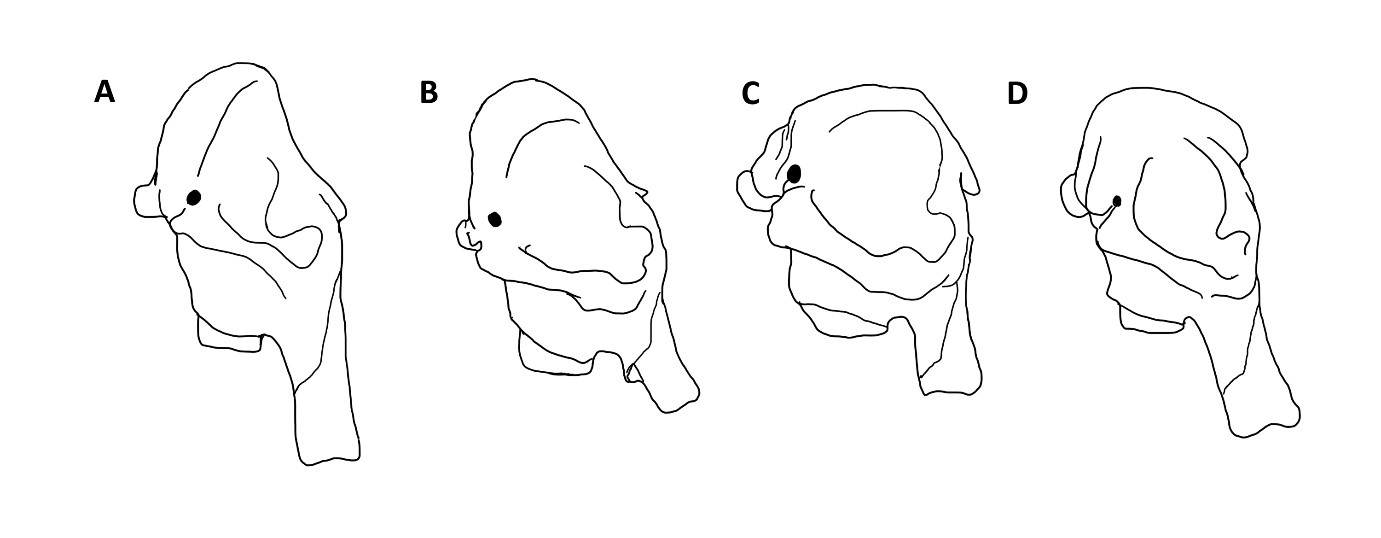
Pracując z materiałem kopalnym najczęściej jesteśmy jednak zdani na różne cechy szkieletu. Jakie cechy szkieletowe wyróżniają więc rodzaj Mammuthus spośród innych słoniowatych? Przy odróżnianiu poszczególnych rodzajów słoni, najważniejsza jest budowa czaszki. Mamuty cechowały się wysklepioną do góry mózgoczaszką (ryc. 2 A, 3 A), podobnie jak słonie azjatyckie (ryc. 2 B, 3 B). Inaczej niż u współczesnych kuzynów, czaszka nie była jednak podzielona na dwa guzy w części szczytowej, ale stanowiła pojedyncze wyniesienie, podobnie, jak u słoni afrykańskich (rodzaj Loxodonta; ryc. 2 C, 3 C). U tych ostatnich, czaszka nie jest jednak wysklepiona. Kości przedszczękowe, czyli te, z których wyrastają słoniom ciosy, były u mamutów ustawione równolegle do siebie, dopiero na końcach rozchylając się lekko na boki (ryc. 2 A). Ta cecha łączy je ze słoniami azjatyckimi (ryc. 2 B), a wyraźnie odróżnia od słoni afrykańskich (ryc. 2 C) oraz kopalnych słoni leśnych (rodzaj Palaeoloxodon; ryc. 2 D), których kości przedszczękowe rozchylały się wachlarzowato na boki. Ciosy mamutów były generalnie długie i grube. Różniły się też nieco strukturą wewnętrzną od ciosów pozostałych słoniowatych. Ciosy mamutów miały również tendencję do skręcania się spiralnie w trakcie wzrostu. U większości innych słoniowatych ciosy zaginają się łukowato w jednej płaszczyźnie, a więc nie skręcają się.
Sytuacja wydaje się być klarowna i odróżnienie mamutów od innych słoniowatych nie powinno nastręczać zbyt wielu trudności. Problem polega jednak na tym, że pomimo swojej wielkości i masy, czaszki słoni są stosunkowo delikatne i bardzo rzadko zachowują się w materiale kopalnym w wystarczająco dobrym stanie. Czasami, takie dobrze zachowane okazy zdarzają się, ale najczęściej znajduje się tylko ich fragmenty, które same w sobie nie dostarczają wszystkich potrzebnych informacji. Tak, jak w przypadku innych ssaków, najczęściej znajdowanymi elementami szkieletu słoni są ich zęby (ryc. 4, 5). Zęby są zbudowane m.in. z zębiny i szkliwa. Obydwie te substancje są bardziej wytrzymałe od kości i przez to zęby mają wyższy potencjał fosylizacyjny, czyli szansę na zachowanie się w dobrym stanie w materiale kopalnym (więcej o zębach ssaków piszemy w poświęconym im artykule).
W historii badań nad kopalnymi ssakami zwykło się przyjmować, że budowa zębów podlega dość ścisłej kontroli genetycznej, a przez to, mogą one służyć jako dobre narzędzie do odróżniania od siebie różnych taksonów, np. rodzajów i gatunków. Dzisiaj wiemy, że wewnętrzna zmienność w obrębie danego taksonu może być całkiem duża. U słoniowatych, w ewolucji uzębienia często dochodziło też do paralelizmu lub konwergencji. Są to sytuacje, w których w dwóch różnych liniach ewolucyjnych dochodzi do wykształcenia bardzo podobnych cech anatomicznych w wyniku działania zbliżonej presji środowiska. Na szczęście, niektóre cechy uzębienia wciąż stanowią rzetelne wskazówki, pozwalające odróżnić od siebie poszczególne rodzaje słoniowatych.
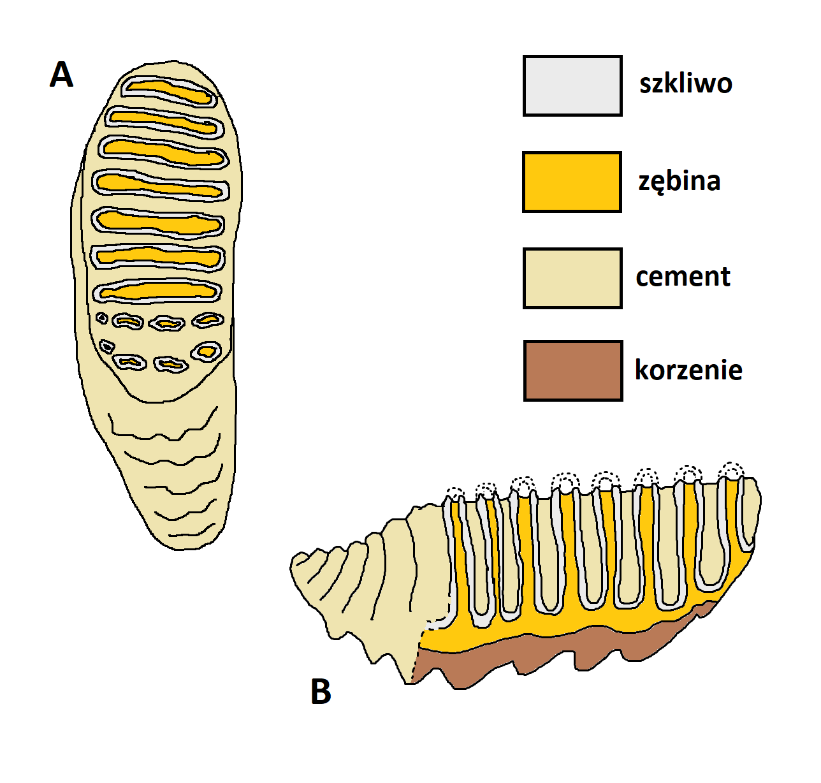
Poza parą ciosów, które są przekształconymi siekaczami, słonie posiadają tylko zęby policzkowe (ryc. 4, 5). Odpowiadają one po kolei drugim mlecznym przedtrzonowcom, trzecim mlecznym przedtrzonowcom, czwartym mlecznym przedtrzonowcom, pierwszym trzonowcom,drugim trzonowcom i trzecim trzonowcom. Zęby te nie występują jednak w pysku jednocześnie, ale wyżynają się po kolei, zastępując siebie nawzajem (opisaliśmy ten proces w artykule poświęconym trąbowcom). W związku z tym, zadanie, które jest rozłożone na kilka zębów w szczękach innych ssaków, u słoni przypada na jeden, ewentualnie dwa zęby. Aby sprostać temu wyzwaniu, zęby policzkowe słoni są zatem bardzo duże. Na pojedynczy ząb składa się seria wysokich, poprzecznych płyt zębowych połączonych ze sobą u podstawy (ryc. 4). Płyty są zbudowane z zębiny, z zewnątrz osłoniętej warstewką szkliwa. Przestrzenie pomiędzy płytami i dokoła nich wypełnione są z kolei cementem – najmniej wytrzymałą z tkanek współtworzących ząb. Tak zbudowana korona jest od spodu zwieńczona licznymi korzeniami.

Płyty zębowe w dopiero co wyżynającym się zębie są na szczycie zwieńczone serią guzków. W czasie pracy, zęby się jednak ścierają, tak że powstają charakterystyczne powierzchnie żujące zbudowane z poprzecznych pierścieni szkliwa (ryc. 4 A, 5). Pierścienie są wewnątrz wypełnione zębiną, a od zewnątrz otoczone cementem. W przeciwieństwie do większości ssaków, słonie nie żują pokarmu poruszając żuchwą na boki. Ruch ten odbywa się w kierunku do przodu i do tyłu. W tym układzie, wstępnie starte zęby działają jak tarki, rozdrabniając skutecznie pokarm roślinny.
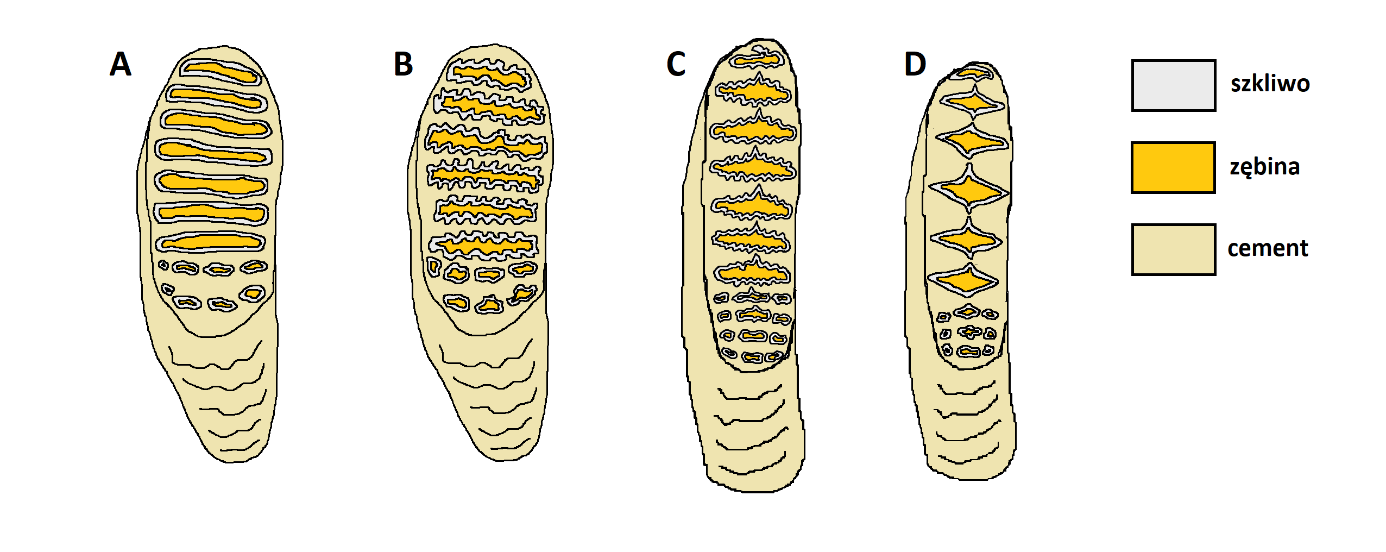
Przy odróżnianiu od siebie poszczególnych słoniowatych, najbardziej wartościowe są trzecie, a zatem ostatnie i największe trzonowce (ryc. 6). Zęby mamutów (ryc. 5, 6 A) najłatwiej odróżnić od zębów słoni afrykańskich (ryc. 6 D). U tych ostatnich, trzonowce są stosunkowo wąskie (w porównaniu z długością) i mają względnie niskie korony. Pierścienie ze szkliwa, które tworzą się w wyniku ścierania zęba, mają charakterystyczny, romboidalny kształt. Samo szkliwo jest stosunkowo grube i słabo pofałdowane. Uważa się, że ten typ uzębienia (nazywany loksodontowym) jest przystosowaniem do odżywiania się głównie pokarmem pochodzącym z roślin drzewiastych (liście, pędy, owoce, gałązki, kora itp.), chociaż współczesne afrykańskie słonie sawannowe (Loxodonta africana) wkomponowują w swoją dietę również trawy.
W porównaniu ze słoniami afrykańskimi, trzonowce mamutów były dość szerokie i wysokie. Pierścienie ze szkliwa miały raczej prostokątny, a nie romboidalny kształt (dłuższe ścianki płyt zębowych były do siebie równoległe), a warstewka szkliwa była cieńsza. Bardzo podobnie zbudowane są też trzonowce słoni z rodzaju Elephas (kopalnych i współczesnych; ryc. 6 B), a do pewnego stopnia również z wymarłego rodzaju Palaeoloxodon (ryc. 6 C). W porównaniu ze słoniami z rodzaju Elephas, mamuty miały bardzo słabo pofałdowaną warstewkę szkliwa, która była też generalnie cieńsza. Rodzaj Palaeoloxodon cechował się stosunkowo wąskimi zębami (tak jak słonie afrykańskie), za to warstewka szkliwa była dość silnie i nieregularnie pofałdowana, a pierścienie szkliwa miały lekko romboidalny kształt – tj. były nieco grubsze po środku niż po bokach (w literaturze, ich kształt często porównuje się do cygara). Rodzaje Mammuthus i Palaeloxodon można również odróżnić po wczesnych etapach ścierania się płyt zębowych. W rodzaju Palaeoloxodon, zanim płyta zetrze się do pojedynczego, poprzecznego pierścienia ze szkliwa, tworzy wzór z trzech mniejszych pętelek, z których środkowa jest wyraźnie bardziej rozciągnięta na boki niż dwie pozostałe. U mamutów i w rodzaju Elephas, temu etapowi odpowiada kilka mniejszych pętelek ze szkliwa, które mają zbliżone rozmiary.

Rodzaje Mammuthus i Elephas są doskonałym przykładem paralelizmu ewolucyjnego w budowie zębów. Rodzaje te można od siebie odróżnić po wyżej opisanych cechach – głównie stopniu pofałdowania warstewki szkliwa oraz jej względnej grubości. Poza tym jednak, w obydwu przypadkach ewolucja przybrała bardzo podobny kierunek. Kolejne, coraz bardziej zaawansowane formy w obydwu rodzajach cechowały się coraz większą ilością, coraz ciaśniej upakowanych płyt zębowych, czemu towarzyszył również wzrost wysokości koron. W obydwu przypadkach, trend ten wyjaśnia się jako adaptację do spożywania coraz większych ilości bardzo włóknistego pokarmu, jakim są trawy. Warto przy tym podkreślić, że o ile zęby zaawansowanych ewolucyjnie mamutów i słoni azjatyckich nadają się dobrze do spożywania traw, to nie oznacza to, że przestały się przez to nadawać do rozdrabniania pokarmu pochodzącego z roślin drzewiastych. Współczesne słonie azjatyckie chętnie jedzą trawy, szczególnie w czasie pory deszczowej, ale rośliny drzewiaste pozostają ważnym elementem ich diety przez cały rok.

Wszystkie rodzaje słoniowatych wywodzą się z Afryki i mamuty nie są tu wyjątkiem. Najstarszym znanym nam przedstawicielem mamutów był Mammuthus subplanifrons (ryc. 7), który żył w południowej i wschodniej Afryce ok. 5-4 mln lat temu (pliocen). Na szczęście, zwierzę to jest znane również z częściowo zachowanych szkieletów. Dzięki temu wiemy, że już wczesne mamuty mogły osiągać całkiem pokaźne rozmiary, nawet 368 cm w kłębie. Ciosy wykazywały już charakterystyczną tendencję do skręcania się. M. subplanifrons posiadał również zespół cech pierwotnych. Jego czaszka była słabo wysklepiona i proporcjonalnie długa. Korony zębów nie były zbyt wysokie, wobec czego zarówno górna szczęka, jak i trzon żuchwy były stosunkowo płytkie (niskie). Żuchwa była też zwieńczona stosunkowo długim rostrum, czyli swojego rodzaju „podbródkiem” (u pierwotniejszych przedstawicieli trąbowców wyrastały z niego dolne ciosy, które uległy zanikowi u zaawansowanych słoniowatych).
Poza pojedynczymi przypadkami, M. subplanifrons znany jest jedynie ze swoich zębów policzkowych i nie ma całkowitej pewności, że wszystkie znaleziska rzeczywiście reprezentują jeden i ten sam gatunek. Początkowa ewolucja słoniowatych jest słabo rozpoznana i tego typu problemy dotyczą również wczesnych przedstawicieli rodzajów Loxodonta, Elephas i Palaeoloxodon. W większości przypadków potrzebne są naukowe rewizje dotychczas dostępnego materiału (głównie zębów), a przede wszystkim konieczne jest odkrycie większej ilości materiału kostnego tych zwierząt (zwłaszcza czaszek).
Kolejnym, domniemanym przedstawicielem afrykańskich mamutów był Mammuthus africanavus sprzed ok. 3-1,65 mln lat. Dla odmiany, jego szczątki znajdowane są głównie w krajach północno-zachodniej Afryki (Maroko, Tunezja, ale także Czad i Libia). Materiał kostny tego gatunku jest jeszcze bardziej skąpy, niż ten M. subplanifrons i również nie ma pewności, czy znajdowane zęby reprezentują jeden gatunek. Jak na mamuta, M. africanavus miał stosunkowo szeroko rozstawione kości przedszczękowe, z których wyrastały słabo skręcające się ciosy. Niektórzy badacze spekulują więc, że tak naprawdę zwierzę reprezentowało rodzaj Loxodonta. Zęby przypisywane M. africanavus pasują jednak swoją morfologią do rodzaju Mammuthus i na chwilę obecną, gatunek ten jest traktowany, jako mamut.
Mamuty przedostały się z Afryki do Eurazji pomiędzy 4 i 3,5 mln lat temu. Ich droga wiodła prawdopodobnie przez obszar lewantu. Z rejonu tego znany jest zespół enigmatycznych skamieniałości, które do tej pory nie zostały dokładnie opisane. Są to głównie zęby różnych słoniowatych, wśród których są też mamuty, których trzecie (diagnostyczne) trzonowce posiadały 8-10 płyt zębowych (ryc. 8). Jest to wartość pośrednia pomiędzy Mammuthus subplanifrons i M. africanavus. Odpowiada ona z kolei najstarszemu gatunkowi mamutów znanemu z Eurazji. Był to Mammuthus rumanus, który żył na terenie Europy i Azji (np. w północnych Chinach) 3,5-2,5 mln lat temu. Niestety, gatunek ten jest znany niemal wyłącznie ze swoich zębów. Tym niemniej, ich charakter pozwala przypuszczać, że rzeczywiście był to mamut. Zęby M. rumanus były ewolucyjnie pierwotniejsze niż dowolnego znanego nauce przedstawiciela Elephas lub Palaeoloxodon (a tylko z tymi rodzajami można by je ewentualnie pomylić). Na chwilę obecną, o ile wiemy, rodzaj Elephas nigdy nie występował w Europie, a jego głównym obszarem dalszej ewolucji (po opuszczeniu Afryki) stała się południowa i południowo-wschodnia Azja. Mamuty natomiast rozwijały się w bardziej północnych obszarach Azji oraz w Europie.
W międzyczasie rozpoczął się plejstocen (ok. 2,58 mln lat temu), czyli tzw. epoka lodowcowa. Zmiany klimatyczne odbywały się początkowo w „krótkich” cyklach, liczących sobie około 41 tys. lat (piszemy o tym w odrębnym artykule) i nie były aż tak drastyczne, jak w późniejszym okresie (pomiędzy 1 mln a 11,7 tys. lat temu). W okresach zimnych (glacjałach), w krajobrazie zaczęło pojawiać się coraz więcej pół-otwartych i otwartych środowisk. W trakcie okresów ciepłych (interglacjałów) Europę bujnie porastały lasy, które również w trakcie ochłodzeń rosły licznie w południowej części kontynentu.
Ok. 2,6 mln lat temu w Europie pojawił się nowy gatunek mamuta – mamut południowy (Mammuthus meridionalis; ryc. 9, 10), który przez krótki czas współistniał z M. rumanus i być może się z nim krzyżował. W końcu jednak nowszy gatunek całkowicie zastąpił starszy. Mamut południowy na szczęście jest nam znany również z materiału szkieletowego i czaszek, a nie tylko z samych zębów (chociaż czaszki są nieliczne). Podobnie jak inne wczesne mamuty, mamut południowy był duży. Byki tego gatunku osiągały przeciętnie 380-420 cm wysokości w kłębie. Zwierzęta miały charakterystycznie opadającą ku tyłowi sylwetkę – jest to cecha często występująca u słoniowatych osiągających bardzo duże rozmiary.
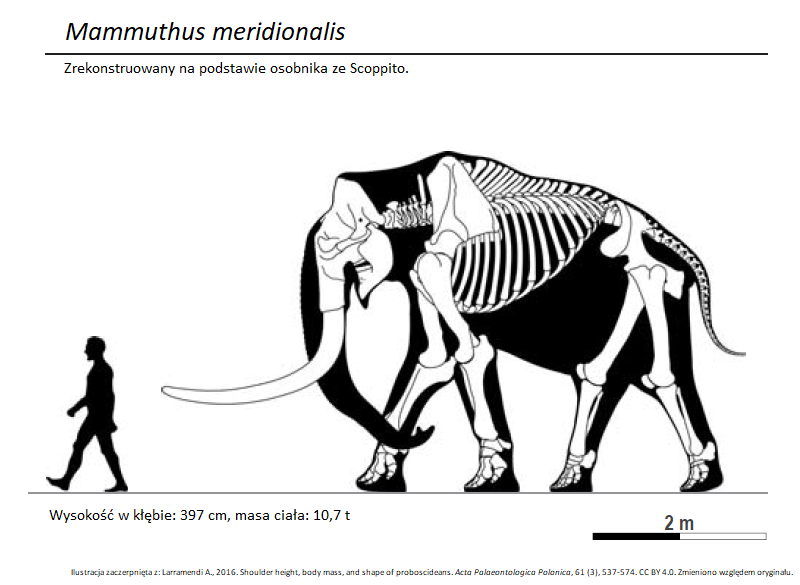
Większość zachowanych czaszek M. meridionalis jest mniej lub bardziej zniekształcona oraz niekompletna. Okazy prezentowane w muzeach były w przeszłości uzupełniane (modelowano brakujące fragmenty), tak że mogą nie odzwierciedlać pierwotnej anatomii. Wydaje się jednak, że istnieją przynajmniej dwa typy budowy. Osobniki z serii typowej z Włoch (tj. te, na podstawie których opisano gatunek) charakteryzują się wyraźnie wklęsłym czołem i silnie wysklepioną czaszką. Osobniki z Francji i Kaukazu posiadają zęby trzonowe, które plasują je w obrębie gatunku M. meridionalis, ale ich czaszki są zbudowane delikatniej, tak że szczyt głowy jest z profilu raczej trójkątny (z bardziej „płaskim” czołem), podobnie jak u późniejszych gatunków mamutów. Pojedynczo, takie czaszki występują również na włoskich stanowiskach. Niektórzy badacze wyróżniają „delikatniejszą” formę, jako odrębny gatunek: Mammuthus gromovi. Forma typowa i M. gromovi bywają też traktowane, jako podgatunki – odpowiednio M. m. meridionalis i M. m. gromovi. Na podstawie różnic w detalach budowy zębów oraz czaszek, w historii badań nad mamutami południowymi wyróżniono jeszcze kilka podgatunków. Niektóre z nich reprezentują różnice pomiędzy regionami (prawdziwe podgatunki), inne – różnice w czasie (chronogatunki), bądź kombinację powyższych. Mamuty południowe posiadały pewne cechy pierwotne. Czaszka była stosunkowo długa, szczęki oraz trzony żuchwy były z kolei relatywnie płytkie (tj. niskie) ponieważ tkwiące w nich zęby nie były jeszcze zbyt wysokie. W trzecich trzonowcach występowało zwykle 12-14 płyt zębowych. Wreszcie, mamuty południowe posiadały wciąż dość długi „podbródek”. Starsze, pierwotniejsze podgatunki (jak M. m. gromovi) miały dłuższe czaszki z „płaskim” czołem oraz niższe zęby z nieco mniejszą liczbą płyt zębowych. Osobniki późne (zaawansowane, jak np. M. m. vestinus, ryc. 9, 10) miały już krótszą, mocno wysklepioną czaszkę (z wklęsłym czołem), a także stosunkowo wysokie zęby.

Zwierzęta posiadały prawdopodobnie mieszaną dietę, obejmującą rośliny drzewiaste, a także trawy i dwuliścienne rośliny zielne. Chętnie zamieszkiwały lasy, chociaż pewnie mogły żerować również na bardziej otwartych terenach (zwłaszcza późniejsze podgatunki). Nauka nie dysponuje żadną formą skamieniałości, która obrazowałaby wygląd tkanek miękkich tego gatunku. Nie wiemy więc, czy mamut południowy okryty był sierścią ani jak duże miał uszy. Ze względu na tryb życia oraz zasiedlane siedliska, można przypuszczać, że okrywa włosowa nie była duża (podobnie, jak u współczesnych słoni), a uszy były być może porównywalne z tymi u słonia azjatyckiego. Taka hipoteza jest oparta na naszej wiedzy o biologii słoni, ale pozostaje oczywiście tylko przypuszczeniem (w świetle braku namacalnych dowodów).

Szczątki mamutów południowych znane są przede wszystkim z Europy, ale ich skamieniałości odkryto również w północnych Chinach. Dowodzi to, że tak, jak jego poprzednik (Mammuthus rumanus), mamut południowy skolonizował znaczną część Eurazji – przynajmniej te rejony, gdzie panował ciepły lub umiarkowany klimat. Mamutom południowym udało się też dostać na Kretę (pomiędzy 2,5-0,8 mln lat temu), gdzie uległy wyspowemu skarleniu, dając początek gatunkowi Mammuthus creticus. Ten mały słoniowaty osiągał zaledwie 113 cm w kłębie i ważył ok. 350 kg. Jego przynależność rodzajowa jest jednak kontrowersyjna. W czasie plejstocenu, na wyspach Morza Śródziemnego pojawiło się kilka gatunków skarlałych słoni. Większości z nich przypisuje się jednak pochodzenie od europejskiego słonia leśnego (Palaeoloxodon antiquus). Gatunek creticus również był tradycyjnie tak postrzegany. Sytuacja zmieniła się wraz z badaniami genetycznymi, które wskazały na przynależność do rodzaju Mammuthus. Część badaczy podważyła wiarygodność otrzymanych wyników, ale jeszcze później, przynależność do rodzaju Mammuthus wykazano na podstawie zębów. Mammuthus creticus był więc prawdopodobnie mamutem, ale trzeba zdawać sobie sprawę z tego, że nie jest to całkiem pewne.
Począwszy od mamuta południowego (Mammuthus meridionalis), dalsza ewolucja mamutów jest w tej chwili całkiem dobrze rozpoznana, chociaż nie tak prosta, jak niegdyś sądzono. Wiąże się ona przede wszystkim ze stopniowym przystosowywaniem się do życia w coraz bardziej suchym, stepowym środowisku, w którym drzewa rosną skąpo lub nie ma ich w ogóle. Głównym źródłem pożywienia stają się wówczas trawy i/lub dwuliścienne rośliny zielne. Tego typu środowisko – step – najwcześniej i w największej skali zaczęło się rozwijać na rozległych obszarach Azji Środkowej (Mongolia, Północne Chiny), znajdujących się w cieniu Himalajów. Ten młody, potężny masyw górski skutecznie blokował dopływ wilgoci w te rejony świata i robi to do dzisiaj.
Pewna subpopulacja mamutów południowych (o „płaskich” czołach) zaczęła zasiedlać obszar Azji Środkowej, adaptując się jednocześnie do napotkanych warunków środowiska. W ten sposób, ok. 2 mln lat temu, powstał nowy gatunek – mamut stepowy (Mammuthus trogontherii; ryc. 11). W porównaniu z mamutem południowym, był on lepiej przystosowany do życia na otwartych terenach trawiastych. Liczba płyt zębowych (w trzecich trzonowcach) wzrosła u niego do 17-22. Zwiększeniu uległa też wysokość koron. Aby pomieścić wyższe zęby, kości szczękowe w czaszce, a także trzony żuchwy, uległy pogłębieniu (zwiększyła się ich wysokość). W ten sposób cała głowa stała się proporcjonalnie wyższa, jednocześnie ulegając skróceniu. Dzięki temu ostatniemu zjawisku, mięśnie obsługujące żuchwę zaczęły pracować w bardziej pionowej linii, co mechanicznie zwiększało ich efektywność. Tradycyjnie uważa się, że zmiany te były adaptacjami do żywienia się trawą, która zawiera proporcjonalnie dużo włókna, a także fitolity (malutkie ziarenka opalu). Czyni to trawę materiałem trudniejszym do przeżucia i bardziej zużywającym zęby, niż pokarm pochodzący od roślin dwuliściennych (zielnych i drzewiastych).
Dzisiaj wiemy, że same właściwości trawy, jako pokarmu, nie usprawiedliwiają tak dalece idących zmian i musiały być one dyktowane również przez inne czynniki, jak np. warunki środowiskowe. Stepy i podobne im, otwarte zbiorowiska trawiaste o suchym klimacie, cechują się dużym zapyleniem powietrza. Pył ten jest pobierany przez roślinożerców wraz z pokarmem. Żywienie się trawami i innymi niskimi roślinami oznacza żerowanie blisko powierzchni gruntu, co również może zwiększać ilość różnych mechanicznych zanieczyszczeń pobieranych razem z pożywieniem. Niewykluczone, że to właśnie te czynniki, a nie same trawy, najmocniej wpływają na ewolucję aparatu gębowego roślinożernych ssaków żyjących w otwartych
Dzisiaj wiemy, że same właściwości trawy, jako pokarmu, nie usprawiedliwiają tak dalece idących zmian i musiały być one dyktowane również przez inne czynniki, jak np. warunki środowiskowe. Stepy i podobne im, otwarte zbiorowiska trawiaste o suchym klimacie, cechują się dużym zapyleniem powietrza. Pył ten jest pobierany przez roślinożerców wraz z pokarmem. Żywienie się trawami i innymi niskimi roślinami oznacza żerowanie blisko powierzchni gruntu, co również może zwiększać ilość różnych mechanicznych zanieczyszczeń pobieranych razem z pożywieniem. Niewykluczone, że to właśnie te czynniki, a nie same trawy, najmocniej wpływają na ewolucję aparatu gębowego roślinożernych ssaków żyjących w otwartych ekosystemach. Szkieletem pozaczaszkowym, mamut stepowy przypominał swojego protoplastę, mamuta południowego. Był duży (byki: 380-420 cm w kłębie), a jego sylwetka opadała ku tyłowi.
Wyodrębnienie się mamuta stepowego nie oznaczało automatycznego końca mamuta południowego. Ten ostatni, w swojej typowej formie (M. meridionalis meridionalis), żył sobie w najlepsze w Europie, przynajmniej do ok. 1,2 mln lat temu. Jak wspomnieliśmy wcześniej, mamut stepowy pojawił się najpierw w środkowej Azji i stamtąd rozprzestrzeniał się na resztę Eurazji (ryc. 12), prawdopodobnie dzieląc się przy tym na kilka podgatunków (nie da się ich jednak wyróżnić po cechach zębów i szkieletu). Około 1,5 mln lat temu dotarł do północno-wschodniej Syberii. Ostatnie, przełomowe badania genetyczne wskazują (DNA liczące sobie ok. 1,2 mln lat!), że na obszarze północno-wschodniej Syberii istniały wówczas dwie niezależne linie ewolucyjne mamutów. Ich zęby są właściwie nieodróżnialne i pasują do charakterystyki mamuta stepowego. Różnice widać dopiero na poziomie genów. Przedstawiciele jednej z tych linii przedostali się ok. 1 mln lat temu do Ameryki Północnej. Wykorzystali w tym celu pomost lądowy, jaki tworzył się w trakcie zlodowaceń pomiędzy północno-wschodnią Syberią a Alaską (tzw. Beringia). Druga z linii ewolucyjnych dała początek m.in. formie, która zawędrowała do Europy.
Do Europy, mamut stepowy (Mammuthus trogontherii) dotarł ok. 1-0,8 mln lat temu. Pokrywa się to w czasie z wydłużeniem się cykli klimatycznych do ok. 100 tys. lat. Zmiany klimatu uległy też zaostrzeniu, tak że w trakcie okresów zimnych (zlodowaceń), na rozległych obszarach Eurazji rozwijał się biom zwany stepo-tundrą lub stepem mamucim. Był to bardzo zimny, unikatowy wariant stepu, o którym piszemy więcej w odrębnym artykule. Stepowienie Eurazji oznaczało redukcję lasów, zamieszkiwanych przez mamuta południowego. Mamut stepowy radził sobie z kolei dobrze zarówno na stepie, jak i w lesie. Okres pomiędzy ok. 800 a 600 tys. lat temu był fazą przejściową. W materiale kopalnym z tego przedziału wiekowego znajdowane są trzonowce, które bądź odpowiadają mamutom południowym (późne, zaawansowane formy), bądź stepowym. Są też pojedyncze znaleziska, które mają cechy pośrednie. Niewykluczone więc, że na granicach zasięgu obydwu gatunków dochodziło do hybrydyzacji – czyli krzyżowania się, czego efektem były właśnie osobniki o pośrednich cechach fizycznych.
Wygląda na to, że ok. 600 tys. lat temu mamut stepowy zastąpił całkowicie mamuta południowego. Niewykluczone, że przysłowiowym „gwoździem do trumny” mamuta południowego było przybycie do Europy słoni leśnych (Palaeoloxodon antiquus), lepiej przystosowanych do leśnego trybu życia. Obydwa gatunki konkurowałyby wówczas o te same zasoby, a z pojedynku zwycięsko wyszedł nowy przybysz (utrzymał potem swoją hegemonię aż do ok. 50-30 tys. lat temu).
W czasie interglacjałów (okresów ciepłych), lasy odzyskiwały tereny utracone podczas zlodowaceń na rzecz stepo-tundry. Tą sytuację wykorzystywały słonie leśne, rozprzestrzeniając się po niemal całej Europie. Mamuty stepowe wycofywały się wówczas w chłodniejsze, stepowe ostoje, gdzie słonie leśne nie czuły się dobrze. Zajmując różne nisze ekologiczne, obydwa gatunki rzadko wchodziły sobie w drogę. Tam gdzie jednak zwierzęta się spotykały, mamuty dominują w zapisie kopalnym.
Przełomowe badania genetyczne, o których już wspomnieliśmy, wskazują, że większość z adaptacji związanych z życiem w chłodnym klimacie, pojawiła się już u syberyjskich mamutów stepowych około 1,2 mln lat temu. Zwierzęta te były więc pokryte futrem i wykształcały podskórną warstwę tłuszczu. Miały też zmodyfikowaną percepcję temperatury i rytm dobowy (zegar biologiczny), dzięki czemu czuły się dobrze w chłodnym klimacie. Nie wiemy, czy adaptacje te były już obecne u najwcześniejszych, środkowo-azjatyckich mamutów stepowych, czy wyewoluowały dopiero w populacji syberyjskiej.
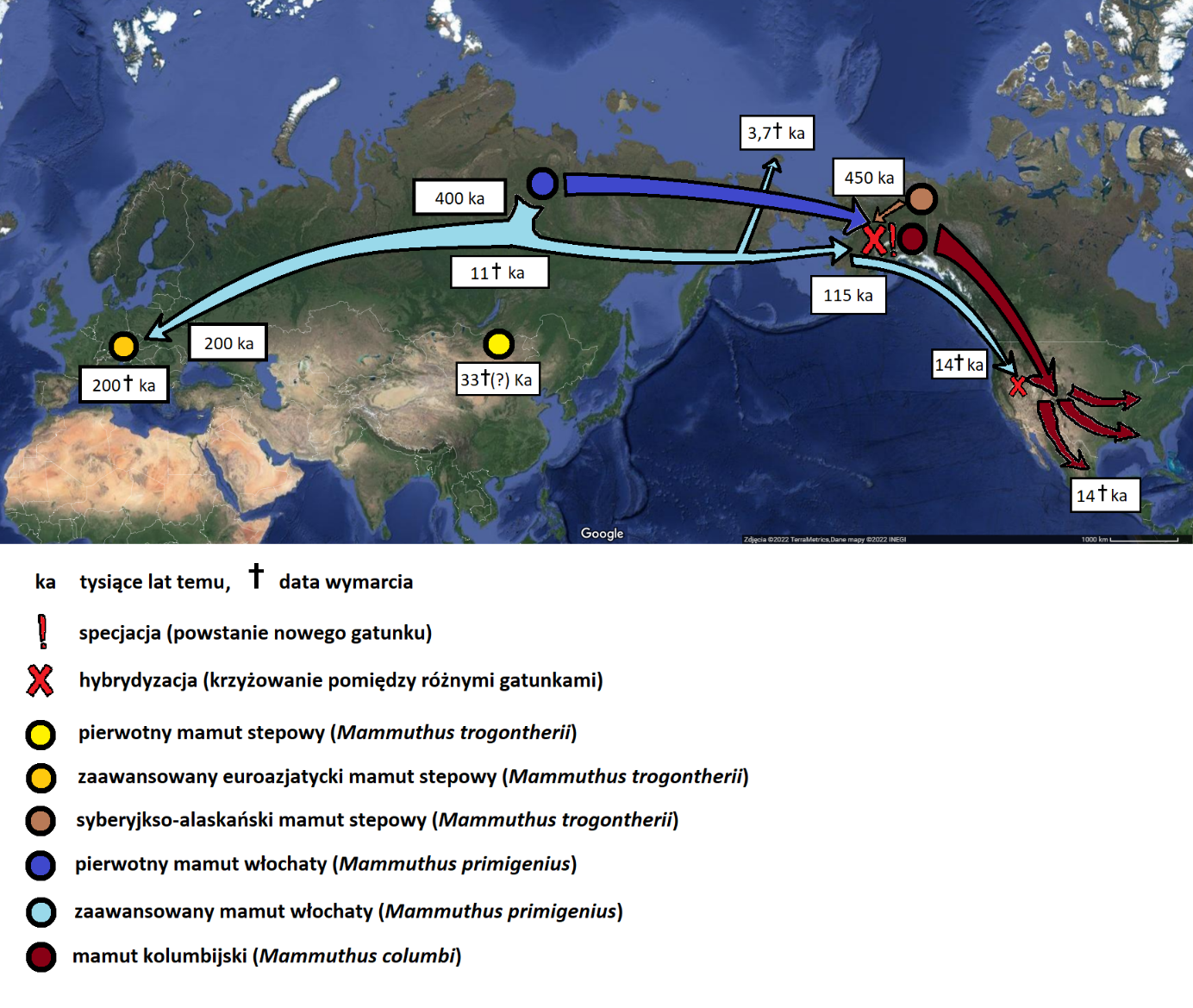
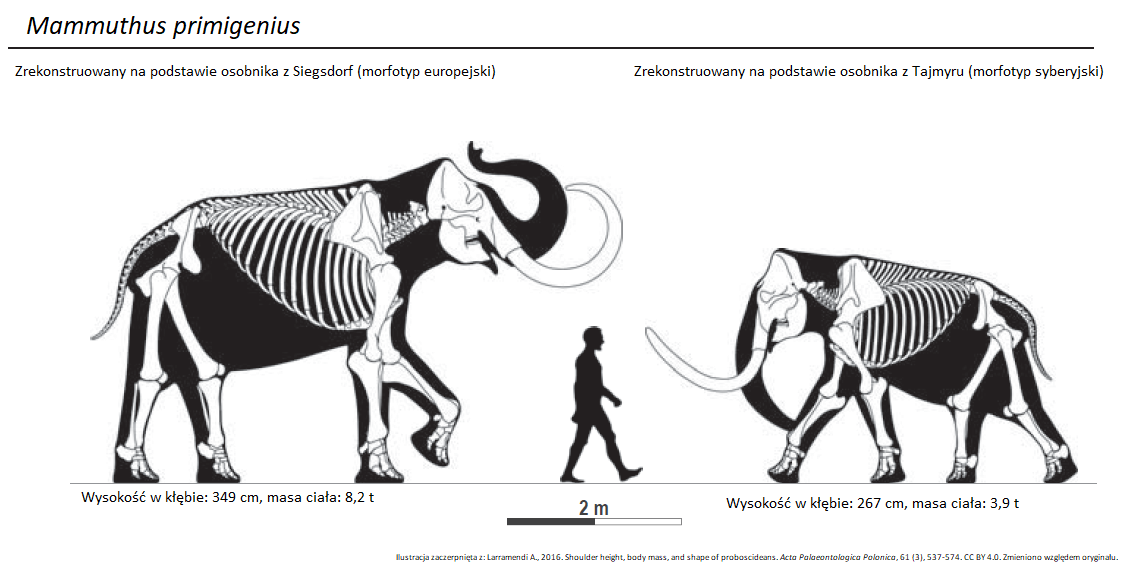
Część z europejskich mamutów stepowych skolonizowała Sardynię (ok. 450 tys. lat temu) i tak, jak wcześniej mamut południowy na Krecie, uległa miniaturyzacji. Efektem był gatunek Mammuthus lamarmorai, który żył na wyspie do około 40 tys. lat temu. Zwierzę osiągało ok. 150 cm w kłębie i ważyło 550-600 kg. W Europie kontynentalnej, mamuty stepowe stały się bardzo konserwatywne ewolucyjnie, tj. niechętnie zmieniały się w czasie. Najistotniejszą zmianą było zmniejszenie przeciętnych rozmiarów ciała w późnym plejstocenie. Przyczyny tego trendu wciąż nie są znane nauce. Na Syberii, zwierzęta zmieniały się jednak dalej. Wśród przedstawicieli tej grupy systematycznie przybywały kolejne płyty zębowe, a czaszka ulegała dalszemu skracaniu. „Podbródek” w żuchwie niemal całkiem zanikł. Liczba płyt zębowych w trzecich trzonowcach zaczęła osiągać rekordowe wartości (24-28). W ten sposób, pomiędzy 700 a 400 tys. lat temu wyodrębnił się mamut włochaty (Mammuthus primigenius; ryc. 13, 16).

Te wczesne formy mamuta włochatego wchodziły w kontakt i krzyżowały się z podgatunkiem mamuta stepowego, który wcześniej przedostał się do Ameryki Północnej (ryc. 13). Zjawisko to zaowocowało (ok. 450 tys. lat temu) powstaniem endemicznego dla Ameryki gatunku, jakim był mamut kolumbijski (Mammuthus columbi; ryc. 15). Dane genetyczne wskazują, że mniej więcej połowa jego genomu pochodziła od wczesnych mamutów włochatych, a druga połowa z amerykańskiej linii mamutów stepowych. Budową szkieletu, mamut kolumbijski najbardziej jednak przypominał mamuta południowego. Był duży (byki: 356-394 cm w kłębie), miał charakterystycznie wklęsły profil czoła oraz opadającą ku tyłowi sylwetkę. Stąd też niegdyś uważano, że to właśnie mamut południowy był przodkiem mamuta kolumbijskiego.Nowsze datowania skamieniałości kazały jednak zrewidować te poglądy i podejrzenie padło wówczas na mamuta stepowego. Wreszcie, wspomniane badania genetyczne przedstawiły nam jeszcze bardziej złożoną historię, obejmującą hybrydyzację.

Mamuty kolumbijskie zachowały zbliżoną do mamuta stepowego liczbę płyt zębowych w trzecich trzonowcach (18-21). Wszystko wskazuje na to, że preferowały też cieplejszy klimat, niż mamut włochaty. Występowały głównie na obszarach obecnych Stanów Zjednoczonych, a nawet dalej na południu, w Meksyku. Nawet w trakcie zlodowaceń, wiele z tych obszarów było przynajmniej częściowo zadrzewionych. Mamut kolumbijski dał początek mamutowi karłowatemu (Mammuthus exilis) – wyspowemu karłowi, który zamieszkiwał Wyspy Santa Barbara (Channel Islands, ang.) u wybrzeży Kalifornii. Budową czaszki, słonik ten mocno przypominał swojego potężnego kuzyna z kontynentu, ale mierzył przeciętnie 175 cm w kłębie i ważył ok. 750 kg. Zaawansowana, syberyjska forma mamuta włochatego (Mammuthus primigenius) cechowała się wyraźnie mniejszymi rozmiarami ciała niż wcześniejszy, syberyjski mamut stepowy. Byki osiągały przeciętnie 266-294 cm w kłębie (ryc. 13, po prawej). Są to rozmiary porównywalne ze współczesnym słoniem azjatyckim. O mamucie włochatym możemy powiedzieć stosunkowo dużo, ponieważ niektóre jego szczątki zachowały się wraz z tkankami miękkimi, zakonserwowane w wieloletniej zmarzlinie Syberii i Alaski (tzw. „mumie”). Dzięki temu wiemy, że zwierzęta te posiadały małe uszy, a ich okrywa włosowa mogła osiągać około metra długości. Pod tym zewnętrznym okryciem znajdował się jeszcze miękki, krótki podszerstek. Futro mamuta kojarzy nam się zwykle z rdzawo-brunatnym zabarwieniem. Częściowo jest to jednak efekt stanu zachowania, a dane genetyczne wskazują, że nie wszystkie mamuty były brązowe i mogły mieć też inne kolory (od czarnego, przez rudy po wersję blond).